
глюкоза
| Структурная формула | |||||||||
|---|---|---|---|---|---|---|---|---|---|

| |||||||||
|
D- глюкоза (слева) и L- глюкоза (справа)
Проекция Фишера , представление открытой цепи | |||||||||
| Общий | |||||||||
| Фамилия | глюкоза | ||||||||
| другие имена |
|
||||||||
| Молекулярная формула | С 6 Н 12 О 6 | ||||||||
| Краткое описание |
D- глюкоза: |
||||||||
| Внешние идентификаторы / базы данных | |||||||||
| |||||||||
| Информация о лекарствах | |||||||||
| Код УВД | |||||||||
| характеристики | |||||||||
| Молярная масса | 180,16 г моль -1 | ||||||||
| Физическое состояние |
фиксированный |
||||||||
| плотность |
1,562 г см −3 |
||||||||
| Температура плавления | |||||||||
| растворимость |
хорошо в воде (470 г л -1 при 20 ° С) |
||||||||
| правила техники безопасности | |||||||||
| |||||||||
| Токсикологические данные | |||||||||
| Насколько это возможно и общепринято, используются единицы СИ . Если не указано иное, приведенные данные относятся к стандартным условиям . | |||||||||
Глюкоза (аббревиатура: Glc ) или глюкоза (от греческого γλυκύς «сладкий» и -оза как суффикс для сахара) - это встречающийся в природе углевод . Из глюкозы энантиомеров , D - глюкоза является естественной формой. Он также известен как глюкоза или пищевые ингредиенты, называемые декстрозой, и является наиболее распространенным моносахаридом (простым сахаром). L- глюкоза может быть представлена синтетически, но не имеет большого значения (объяснение обозначений « D » и « L » см. В проекции Фишера ). Если глюкоза указана без какого-либо дополнительного названия ( префикса ), имеется в виду D- глюкоза.
В живых организмах глюкоза является основным источником энергии. Он хранится в виде полимерного глюкана , у животных - в виде гликогена, а у растений - в виде крахмала . Как компонент целлюлозы , он является основным компонентом стенок растительных клеток и, следовательно, наиболее распространенным углеводом. Глюкоза используется в медицинских инфузионных растворах и считается основным лекарством .
история
Глюкоза была впервые выделена из изюма Андреасом Сигизмундом Маргграфом в 1747 году . Он был выделен из винограда Иоганном Тобиасом Ловицем в 1792 году и признан отличным от тростникового сахара ( сахарозы ). Глюкоза - это термин, придуманный Жаном Батистом Дюма в 1838 году и закрепившийся в химической литературе. По Фридрих Кекуле декстрозы предложения приходит (от латинского Dexter = справа), как глюкоза в водном растворе плоскость линейно поляризованный свет на правильные повороты. В противоположности этому , D - фруктоза (а кетогексоз ) и L - глюкоза поворот линейно поляризованный свет влево . Ранняя номенклатура d и l, основанная на этом, была оставлена в пользу обозначения D и L , которое относится к абсолютной конфигурации асимметричного центра, наиболее удаленного от карбонильной группы, и той, что соответствует D или L -глицеральдегиду.
Структура глюкозы и структурные отношения с другими моносахаридами были описаны Эмилем Фишером между 1891 и 1894 годами с помощью проекции Фишера и стали важной вехой в химии природных продуктов , за что он получил Нобелевскую премию по химии в 1902 году . Его первый успешный синтез глюкозы подтвердил теории Якоба Хенрикуса ван 'т Гоффа о тетраэдрическом расположении химических связей в органических углеродных соединениях и о химической кинетике . Полученное таким образом знание дало возможность ввести систематическую номенклатуру для стереоизомерных углеводов с использованием обычных имен со ссылкой на их пространственную симметрию (например , Фишер номенклатура, D / L номенклатура).
Отто Мейерхоф получил Нобелевскую премию по физиологии и медицине в 1922 году за открытие метаболизма глюкозы . Ганс фон Эйлер-Челпин вместе с Артуром Харденом получил Нобелевскую премию по химии в 1929 году за свои исследования «ферментации сахара и того, как ферменты способствуют этому процессу». В 1947 году Карл и Герти Кори получили Нобелевскую премию по физиологии и медицине за открытие рециркуляции молочной кислоты, вырабатываемой в мышцах в результате гликолиза, для глюконеогенеза и синтеза гликогена в печени, а Бернардо Хусей за открытие роли гормонов гипофиза. в метаболизме глюкозы. Луис Лелуар получил Нобелевскую премию по химии в 1970 году за открытие нуклеотидов сахара, образующихся из глюкозы в процессе биосинтеза углеводов .
характеристики
Глюкоза, как правило , в твердой форме в виде моно- гидрат закрыт пирано кольцо перед ( Hydratdextrose ). В водном растворе, с другой стороны, он имеет открытую цепь в небольшой степени и преимущественно находится в форме α- или β-пиранозы, некоторые из которых сливаются друг с другом посредством мутаротации. Три известные формы могут быть кристаллизованы из водных растворов: α-глюкопираноза, β-глюкопираноза и гидрат β-глюкопиранозы. Глюкоза является компонентом двойного сахара, лактозы (молочный сахар), сахарозы (тростниковый или свекольный сахар) и мальтозы, а также нескольких сахаров, таких как рафиноза, и нескольких сахаров, таких как крахмал и амилопектин , гликоген и целлюлоза . Температура стеклования глюкозы составляет 31 ° C, а постоянная Гордона-Тейлора (экспериментально определенная константа для предсказания температуры стеклования при различных массовых долях смеси двух веществ) составляет 4,5.
Глюкоза, вероятно, является наиболее распространенным природным моносахаридом, потому что она меньше реагирует с белками посредством гликирования, чем другие моносахариды. Другая гипотеза состоит в том, что с глюкозой в форме β- D- глюкозы в качестве единственной D- альдогексозы все пять гидроксизаместителей находятся в экваториальном положении и поэтому более доступны для химических реакций, например, для этерификации или образования ацеталей. По этой причине D- глюкоза также является наиболее предпочтительным строительным блоком природных полисахаридов (гликанов).
Систематика глюкозы
| Сравнение различных форм и представлений D- глюкозы | ||
|---|---|---|
| Формула клина | Обозначение Хаворта | |

|
 α- D- глюкофураноза |
 β- D- глюкофураноза |
 α- D- глюкопираноза |
 β- D- глюкопираноза |
|
| α- D- глюкопираноза в (1) Толленс / Фишер (2) Хаворт (3) кресло-репрезентация (4) стереохимическая картина | ||

| ||
Представление α-, показанного выше, D- глюкозы в виде α- D- глюкопиранозы, см. 2 , кроме того, вызвано ее проекцией Хаворта и нарисовано новое кольцо, образованное на этой иллюстрации для удобства как (что может не соответствовать действительности, поскольку много целей, но этого достаточно). В результате замыкания кольца первый атом углерода становится новым, дополнительным хиральным центром , в результате чего становится возможной пара аномеров - стереоизомерные сахара, которые различаются только конфигурацией в их новообразованном хиральном центре во время замыкания кольца и, следовательно, особый случай Эпимеры .
Структура, в которой вновь созданная группа ОН полуацеталя указывает «вниз» в своей проекции Хаворта (в осевом направлении), называется α- D- глюкозой, а группа ОН (экваториальная) «направлена вверх» как β- D - глюкоза. В общем, в случае α-аномерных сахаров группа ОН, вновь образованная при замыкании кольца, находится в проекции Хаворта на стороне плоскости кольца, противоположной концевой группе СН 2 ОН, тогда как в форме β она находится на той же стороне. боковая сторона. Между прочим, это определение также применяется таким же образом ко всем другим сахарам D и L, а также - при условии, что здесь нет концевой группы СН 2 ОН, такой как, например, Б. во фруктопиранозе , часть замыкания кольца - как для альдоз, так и для кетозов.
Проекция Фишера сбивает с толку циклические полуацетальные формы, см. 1 . Чтобы уточнить угловое расположение углеродной цепи, выбрано кресло 3 . Представление 4 также является обычным и стереохимически однозначным.
Поведение в водном растворе

Кольцо можно открывать и закрывать в водном растворе, так что существует равновесие между пиранозой - (шестичленное кольцо с эндоциклическим атомом кислорода, 99,75%), формой фуранозы (пятичленное кольцо, в следах) и формой альдегида с открытой цепью. (0,25%). Добавление кислоты или щелока ускоряет этот процесс. Поскольку во время замыкания кольца может возникнуть альфа- или бета-форма, существует также баланс между альфа-формой (36%) и бета-формой (63,9%). Баланс есть, как можно видеть на проценты, на стороне бета- D - глюкозы. Это более стабильный аномер , поскольку все гидроксильные группы в конформации кресло расположены экваториально и, таким образом, находятся на максимально возможном расстоянии друг от друга. Тот факт, что 36% α-аномера все еще присутствует, несмотря на его аксиальную ОН-группу, указывает на дальнейшее влияние ( аномерный эффект ). Без аномерного эффекта было бы соотношение 11% α-формы к 89% β-формы.
Мутаротация
Преобразование между двумя аномерами может наблюдаться в поляриметре, потому что чистая α- D- глюкоза с удельным углом вращения + 112,2 ° · мл · дм -1 · г -1 , имеет чистую β- D- глюкозу +17 , 5 ° мл дм −1 г −1 . Если через определенное время установилось равновесие, угол поворота составляет + 52,7 ° · мл · дм −1 · г −1 . Это изменение угла поворота известно как мутаротация . Это преобразование можно значительно ускорить , добавив кислоту или основание . Равновесие устанавливается через альдегидную форму с открытой цепью.

Изомеризация
В разбавленном растворе гидроксида натрия манноза , глюкоза и фруктоза превращаются друг в друга ( перегруппировка Лобри-де-Брюна-Альберда-ван-Экенштейна ), так что между этими изомерами устанавливается равновесие . Реакция протекает через ендиол :

Вспомогательные средства для стереохимии

Для того , чтобы запомнить конфигурацию глюкозы в проекции Фишера, существует следующая осел мост : Позиции гидроксильных групп справа и слева от углеродной цепи может быть «символом» с помощью звукоподражания из звука мартеновского Горна («Та-Тю-Та-Та»). С другой стороны, расположение гидроксильных групп в галактозе можно запомнить как синий свет (см. Иллюстрацию).
Вы можете использовать слово «HochBett» (B для бета), чтобы запомнить, какая форма глюкозы присутствует. Если гидроксильная группа C-1 находится наверху («высоко») в проекции Хауорта, то это бета-форма («слой»).
Вхождение
Глюкоза присутствует во всех живых существах, потому что это центральный компонент метаболизма всех живых существ.
| еда | Всего углеводов, включая клетчатку |
Общий сахар | Фруктоза | глюкоза | Сахароза | Соотношение
фруктоза / глюкоза |
Сахароза в% от общего сахара |
|---|---|---|---|---|---|---|---|
| фрукты | |||||||
| яблоко | 13,8 | 10,4 | 5.9 | 2,4 | 2.1 | 2.0 | 19,9 |
| абрикос | 11.1 | 9.2 | 0,9 | 2,4 | 5.9 | 0,7 | 63,5 |
| банан | 22,8 | 12.2 | 4.9 | 5.0 | 2,4 | 1.0 | 20,0 |
| Инжир сушеный | 63,9 | 47,9 | 22,9 | 24,8 | 0,9 | 0,93 | 0,15 |
| Виноград | 18,1 | 15.5 | 8.1 | 7.2 | 0,2 | 1.1 | 1 |
| Пуповина апельсин | 12,5 | 8,5 | 2,25 | 2.0 | 4.3 | 1.1 | 50,4 |
| персик | 9,5 | 8,4 | 1.5 | 2.0 | 4.8 | 0,9 | 56,7 |
| груша | 15.5 | 9,8 | 6.2 | 2,8 | 0,8 | 2.1 | 8.0 |
| ананас | 13,1 | 9.9 | 2.1 | 1,7 | 6.0 | 1.1 | 60,8 |
| слива | 11,4 | 9.9 | 3.1 | 5.1 | 1.6 | 0,66 | 16.2 |
| овощи | |||||||
| Свекла | 9,6 | 6,8 | 0,1 | 0,1 | 6.5 | 1.0 | 96,2 |
| морковь | 9,6 | 4,7 | 0,6 | 0,6 | 3,6 | 1.0 | 77 |
| перец | 6.0 | 4.2 | 2.3 | 1.9 | 0,0 | 1.2 | 0,0 |
| лук | 7,6 | 5.0 | 2.0 | 2.3 | 0,7 | 0,9 | 14,3 |
| сладкая картошка | 20,1 | 4.2 | 0,7 | 1.0 | 2,5 | 0,9 | 60,3 |
| сладкий картофель | 27,9 | 0,5 | следы | следы | следы | - | следы |
| Сладкая палочка | 13–18 | 0,2-1,0 | 0,2-1,0 | 11–16 | 1.0 | высокий | |
| сахарная свекла | 17–18 | 0,1-0,5 | 0,1-0,5 | 16-17 | 1.0 | высокий | |
| зерно | |||||||
| Кукуруза | 19.0 | 6.2 | 1.9 | 3,4 | 0,9 | 0,61 | 15.0 |
Индустриальное производство

В промышленности глюкозу получают из крахмала путем ферментативного гидролиза с использованием глюкозоамилазы или с использованием кислот , причем ферментативный гидролиз в значительной степени заменил гидролиз, катализируемый кислотой. Это создает сироп глюкозы (ферментативно с более чем 90% глюкозы в сухом веществе) с годовым объемом производства в мире 20 миллионов тонн (по состоянию на 2011 год). Именно здесь раньше использовалось название «крахмальный сахар». В качестве источников крахмала используются кукуруза , картофель , рис , пшеница , рожь , маниока , сладкий картофель и саго . Амилазы в основном происходят из Bacillus licheniformis или Bacillus subtilis (штамм MN-385), которые более термостабильны, чем ферменты, используемые ранее. Начиная с 1982 г. пуллуланазы из Aspergillus niger использовались в производстве сиропа глюкозы для преобразования амилопектина в амилозу, тем самым увеличивая выход глюкозы. Реакцию проводят при pH 4,6-5,2 и температуре 55-60 ° C. Кукурузный сироп содержит от 20% до 95% глюкозы в сухом веществе. Японская форма глюкозного сиропа Mizuame производится из сладкого картофеля или рисового крахмала. Мальтодекстрин содержит около 20% глюкозы.
Превращение во фруктозу
В США кукуруза используется почти исключительно с помощью амилазы и изомеразы глюкозы для производства изоглюкозы пищевой сладости , которая представляет собой смесь глюкозы и фруктозы (также кукурузный сироп HFCS с высоким содержанием фруктозы ). Фруктоза имеет более высокую сладость, чем глюкоза, с той же физиологической теплотворной способностью 374 килокалорий на 100 г. Ежегодное мировое производство изоглюкозы составляет восемь миллионов тонн (по состоянию на 2011 год).
Промышленное использование
Прежде всего, глюкоза используется для производства фруктозы и продуктов, содержащих глюкозу . В пище он используется как подсластитель , как увлажнитель , для увеличения объема и создания более мягкого ощущения во рту . Различные источники глюкозы, такие как виноградный сок (для вина) или солод (для пива), используются для ферментации в этанол в процессе производства алкогольных напитков . В большинстве безалкогольных напитков в США используется HFCS-55 (то есть 55% фруктозы), в то время как в большинстве других подслащенных HFCS пищевых продуктов в США используется HFCS-42. В соседней Мексике , с другой стороны, тростниковый сахар используется в качестве подсластителя в безалкогольном напитке Coca-Cola , который имеет более высокую подслащивающую способность. Сироп глюкозы также используется в производстве сладостей, таких как конфеты , ирис и помадка . Типичные химические реакции глюкозы в сухих приготовления пищи методов являются карамелизациями , и с аминокислотами, в реакции Майяра .
Глюкоза подходит в качестве субстрата для промышленной ферментации , например, с Clostridium thermoaceticum для производства уксусной кислоты , с Penicillium notatum для производства арабоаскорбиновой кислоты , с Rhizopus delemar для производства фумаровой кислоты , с Aspergillus niger для производства глюконовой кислоты. кислоты , с Candida brumptii для производства изоазонной кислоты , с Aspergillus terreus для производства итаконовой кислоты , с Pseudomonas fluorescens для производства 2-кетоглюконовой кислоты , с Gluconobacter suboxydans для получения 5-кетоглюконовой кислоты , с Aspergillus oryzae для приготовления койевых кислот с Lactobacillus delbruckii для приготовления молочной кислоты , с Lactobacillus BREVIS для приготовления яблочной кислоты , с Propionibacter shermanii для получения пропионовой кислоты , с синегнойной палочкой для получения пировиноградной кислоты и с Gluconobacter suboxydans для получения винной кислоты .
В следующей таблице дается краткий обзор важных продуктов, которые можно производить биотехнологически. Жирным шрифтом выделены промышленно интересные продукты и их предварительные стадии :
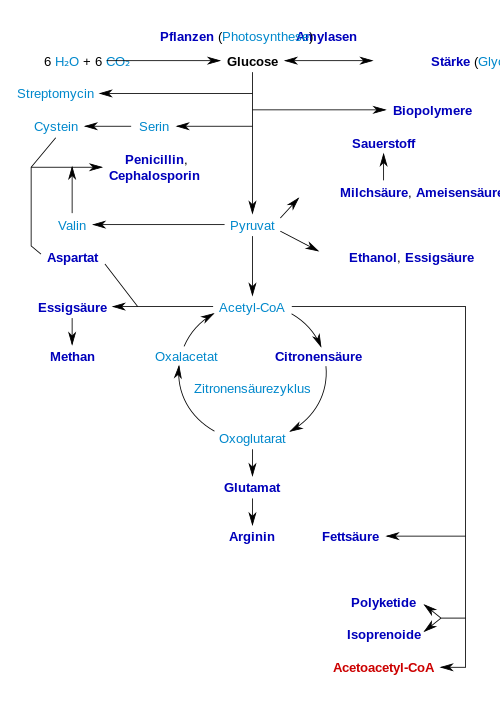

Фермент целлюлаза используется для производства глюкозы из целлюлозы для производства этанола ( целлюлоза-этанол ) для использования в качестве биотоплива .
биохимия
Глюкоза в основном вырабатывается растениями с помощью фотосинтеза из солнечного света, воды и углекислого газа и может использоваться всеми живыми существами в качестве источника энергии и углерода. Большая часть глюкозы в растениях и животных не свободна, а находится в связанной форме, например, в форме лактозы или свекольного сахара (сахарозы) , или в форме полимеров, таких как крахмал или целлюлоза, которые являются резервными веществами или компоненты клеточной стенки растений . Эти полимеры сначала расщепляются на глюкозу с помощью ферментов, когда их едят животные, грибы и бактерии . У людей это иногда случается при жевании с помощью амилазы, содержащейся в слюне , и с помощью мальтазы . Все живые существа также могут сами производить глюкозу из определенных исходных материалов, если возникает необходимость. Нервные клетки , клетки мозгового вещества почек и эритроциты зависят от глюкозы для производства энергии. У взрослого человека содержится около 18 г глюкозы, из которых около 4 г находится в крови. Примерно 180–220 г глюкозы образуется в печени взрослого человека за 24 часа.
Предшественники других биомолекул
У живых существ глюкоза превращается в несколько других химических соединений, которые являются исходным продуктом различных метаболических путей. Все другие моносахариды, такие как фруктоза (через полиольный путь ), манноза (эпимер в положении 2), галактоза (эпимер в положении 4), фукоза , различные уроновые кислоты и аминосахара производятся из глюкозы . В дополнение к фосфорилированию до глюкозо-6-фосфата , которое является частью гликолиза , глюкоза может первоначально окисляться до глюконо-1,5-лактона, когда она расщепляется. У некоторых бактерий глюкоза служит строительным блоком в биосинтезе трегалозы или декстрана, а у животных - строительным блоком для гликогена. Глюкоза также может превращаться во фруктозу с помощью бактериальной ксилозоизомеразы . Кроме того, все заменимые аминокислоты , сахарные спирты, такие как маннит и сорбит , жирные кислоты , холестерин и нуклеиновые кислоты, производятся из продуктов метаболизма глюкозы . Наконец, глюкоза используется в качестве строительного блока в гликозилирования к гликопротеинам , необходимым для функции многих белков , а также в других гликозилированных веществах , такие как гликолипиды , пептидогликаны и гликозиды . Гликозилирование катализируется гликозилтрансферазами и может снова отщепляться гликозидазами .
запись
У людей глюкоза, поступающая с пищей, сначала связывается с рецептором сладкого вкуса на языке. Этот комплекс белков T1R2 и T1R3 позволяет идентифицировать источники пищи, содержащие глюкозу. Глюкоза поступает в основном с пищей - около 300 г в день вырабатывается путем преобразования пищи, но она также синтезируется из других метаболитов в клетках организма. Глюкоза входит в состав многих углеводов и может отделяться от них с помощью определенных ферментов . Глюкозидазы (подгруппа гликозидаз) сначала катализируют гидролиз длинноцепочечных глюкозосодержащих полисахаридов с удалением концевой глюкозы. Дисахариды, в свою очередь, в основном расщепляются на глюкозу специальными гликозидазами . Названия разрушающих ферментов часто происходят от соответствующих поли- и дисахаридов; Например, есть амилазы (из амилозы , входящей в состав крахмала ), целлюлазы (из целлюлозы ), хитиназы (из хитина ) и другие для разрушения полисахаридных цепей ; кроме того, для расщепления дисахаридов используются лактаза , сахараза , трегалаза и другие. У человека известно около 70 генов , кодирующих гликозидазы. Они выполняют функции пищеварения и расщепления гликогена, сфинголипидов , мукополисахаридов и поли (АДФ-рибозы) . В организме человека, полисахариды , содержащие глюкозу, частично разбиты во время жевания с помощью амилазы , который содержится в слюне , а также с помощью мальтаза , лактазы и сахараза в щеточной каемке в тонкой кишке .
Чтобы попасть в клетки и компартменты клеток через клеточную мембрану или выйти из них, глюкоза нуждается в специальных транспортных белках из суперсемейства основных посредников . В тонком кишечнике (точнее в тощей кишке ), глюкоза поглощаются в эпителиальные клетки кишечника с помощью транспортера глюкозы через вторичный активный транспорт механизм называется ионно-глюкоза натрия symport ( натрий / глюкоза котранспортер 1 ) . Он передается на базолатеральную сторону кишечных эпителиальных клеток через переносчик глюкозы GLUT2 , так же как и его захват гепатоцитами , клетками почек , β-клетками островков Лангерганса, нейронами, астроцитами и таницитами . Глюкоза попадает в печень через воротную вену и хранится там в виде гликогена. В клетке печени он фосфорилируется до глюкозо-6-фосфата глюкокиназой в положении 6 ; поэтому она больше не может выходить из камеры. С помощью глюкозо-6-фосфатазы глюкозо-6-фосфат при необходимости превращается обратно в глюкозу исключительно в печени, так что он доступен для поддержания достаточной концентрации в крови. В других клетках захват происходит посредством пассивного транспорта через один из 14 белков GLUT. В других типах клеток фосфорилирование осуществляется гексокиназой , после чего глюкоза больше не может диффундировать из клетки.
GLUT1 образуется большинством типов клеток и имеет особое значение для нервных клеток, а поджелудочная железа содержит β-клетки . GLUT3 часто образуется нервными клетками. Глюкоза из кровотоке абсорбируется на мышечные клетки (скелетных мышц и сердечной мышцы) и жировые клетки через GLUT4 . GLUT14 производится только в семенниках . Избыточная проглоченная глюкоза расщепляется и превращается в жирные кислоты, которые хранятся в виде триацилглицеридов . В почках глюкоза абсорбируется из мочи через SGLT1 и SGLT2 в апикальных клеточных мембранах и передается через GLUT2 в базолатеральные клеточные мембраны. Около 90% абсорбции глюкозы почками происходит через SGLT2 и около 3% через SGLT1.
биосинтез
Метаболический путь образования глюкозы из небольших молекул с двумя-четырьмя атомами углерода, который заканчивается молекулой глюкозы с шестью атомами углерода, называется глюконеогенезом и происходит во всех живых существах. Посредством глюконеогенеза организм может накапливать глюкозу из других продуктов метаболизма, включая лактат или определенные аминокислоты, потребляя при этом энергию. Меньшие исходные материалы являются результатом других метаболических путей, а у растений, в конечном итоге, возникают в результате ассимиляции углекислого газа . В конечном итоге почти все биомолекулы образуются в результате ассимиляции углекислого газа растениями во время фотосинтеза. Свободная энергия образования & alpha ; D - глюкозы составляет 917,2 кДж на моль. В организме человека глюконеогенез происходит в печени и почках , но также и в других типах клеток . Трубчатые клетки почек также могут вырабатывать глюкозу.
Распад гликогена известен как гликогенолиз, распад крахмала как распад крахмала. Около 150 г гликогена хранится в печени и около 250 г в скелетных мышцах . Глюкоза, высвобождаемая в мышечных клетках при расщеплении гликогена, не может попадать в кровоток, однако, поскольку глюкоза фосфорилируется гексокиназой, глюкозо-6-фосфатаза не образуется для удаления фосфатной группы и глюкозо-6-фосфата в организме. в отличие от глюкозы транспортного белка не существует.
Разборка
Глюкоза расщепляется в процессе метаболизма посредством гликолиза и пентозофосфатного пути. Гликолиз используется всеми живыми организмами с небольшими вариациями, и все организмы вырабатывают энергию за счет распада моносахаридов. В ходе дальнейшего метаболизма он может полностью распадаться на воду и диоксид углерода посредством окислительного декарбоксилирования , цикла лимонной кислоты и дыхательной цепи . При недостатке кислорода у животных происходит анаэробное разложение глюкозы до лактата путем молочнокислого брожения и затрачивается меньше энергии. У млекопитающих лактат из мышц через кровоток достигает печени, где происходит глюконеогенез ( цикл Кори ). Другие формы брожения также встречаются у других живых существ . При большом количестве глюкозы метаболит ацетил-КоА также используется для синтеза жирных кислот . Глюкоза также пополняет запасы гликогена в организме, которые в основном находятся в печени и скелетных мышцах. Эти процессы регулируются гормонально.

Бактерия Escherichia coli может расти на питательных средах , в которых глюкоза является единственным источником углерода. У некоторых бактерий и - в модифицированной форме - также у архей глюкоза расщепляется по пути Энтнера-Дудорова .
Опухолевые клетки часто растут сравнительно быстро и потребляют за счет гликолиза количество глюкозы выше среднего, что приводит к образованию лактата, конечного продукта ферментации у млекопитающих, даже в присутствии кислорода . Этот эффект известен как эффект Варбурга . Различные SGLT и GLUT все чаще образуются для повышенного поглощения глюкозы опухолями . В дрожжах при высоких концентрациях глюкозы, даже в присутствии кислорода (обычно приводит к дыханию , но не к ферментации), этанол образуется путем ферментации. Этот эффект известен как эффект Крэбтри .
Теплотворная способность

Физиологическое теплотворная глюкозы, в зависимости от источника, 16,2 кДж на грамм или 15,7 кДж / г (3,74 ккал / г). Высокая доступность углеводов через растительную биомассу привела к появлению множества методов в ходе эволюции , особенно микроорганизмов , для использования энергии и глюкозы, запасающей углерод. Существуют различия в конечном продукте, из которого получается конечный продукт, который больше не может использоваться для выработки энергии. Здесь присутствие отдельных генов и их генных продуктов, ферментов, решает, какие реакции возможны (см. Рисунок). Метаболический путь гликолиза используется почти во всех живых существах. Существенное отличие в этом случае является производством НАДФА в качестве восстановителя для анаболизма , которые в противном случае пришлось бы производить косвенно.
Концентрации глюкозы
Уровень глюкозы в крови называется сахаром в крови . Уровень сахара в крови регулируется связывающими глюкозу нервными клетками гипоталамуса . Кроме того, глюкоза в головном мозге связывается с рецепторами глюкозы в центре вознаграждения в прилежащем ядре . Связывание глюкозы с рецептором сладкого вкуса на языке вызывает высвобождение различных гормонов энергетического метаболизма с приемом глюкозы и без нее, что приводит к увеличению поглощения клетками и снижению уровня сахара в крови. С другой стороны, подсластители не снижают уровень сахара в крови.
У здорового человека уровень сахара в крови натощак, т.е. ЧАС. после ночного голодания примерно от 70 до 100 мг / дл крови (от 4 до 5,5 мМ ). Эти значения , измеренные в плазме крови составляет около 10-15% выше. Кроме того, значения в артериальной крови выше, чем концентрации в венозной крови, поскольку глюкоза всасывается в ткань во время прохождения через капиллярное русло. В капиллярной крови, которая часто используется для определения уровня сахара в крови, значения иногда выше, чем в венозной крови. Содержание глюкозы в крови регулируется гормонами инсулином , инкретином и глюкагоном : инсулин снижает уровень глюкозы, глюкагон повышает его. Кроме того, гормоны адреналин , тироксин , глюкокортикоиды , соматотропин и адренокортикотропин приводят к повышению уровня глюкозы. Существует также гормонально-независимая регуляция, называемая ауторегуляцией глюкозы . После еды концентрация сахара в крови увеличивается. Значения выше 180 мг / дл в цельной венозной крови являются патологическими и называются гипергликемией , значения ниже 40 мг / дл называются гипогликемией .
При использовании глюкоза высвобождается в кровоток глюкозо-6-фосфатазой из глюкозо-6-фосфата, а также из гликогена из печени и почек, посредством чего достигается гомеостаз концентрации глюкозы в крови. У жвачных животных концентрация глюкозы в крови низкая (60 мг / дл у крупного рогатого скота и 40 мг / дл у овец), потому что углеводы кишечной флоры в большей степени преобразуются в жирные кислоты с короткой цепью .
В мозге , который полагается на глюкозу в качестве основного поставщика энергии , концентрация глюкозы обычно составляет от 4 до 6 мМ (5 мМ соответствует 90 мг / дл), но снижается до 2–3 мМ во время голодания . Низкие концентрации глюкозы в головном мозге отрицательно влияют на способность думать, самоконтроль и волю . Ниже 1 мМ возникает путаница, а ниже 1 мМ - кома .
Гликемический индекс является показателем скорости поглощения и превращения в глюкозу крови из углеводов и глотавших определяются как интеграл от уровня глюкозы в крови после приема пищи по сравнению с глюкозой (заглатывание глюкозы определяются как 100). Клиническое значение гликемического индекса является спорным, поскольку продукты с высоким содержанием жира замедляют всасывание глюкозы и, таким образом, снижают гликемический индекс, например, мороженое. Альтернативным показателем для решения той же проблемы является индекс инсулина , измеряемый как влияние потребления углеводов на уровень инсулина в крови. Гликемической нагрузки является показателем количества глюкозы добавляют к крови после приема углеводов и основан на гликемический индекс и количество пищи , съеденной.
Патобиохимия
Аутоиммунный диабет
В ходе проведения аутоиммунной реакции против бета - клеток в островках из Лангерганса в тех поджелудочной железы , что бета - клетки не погибают, а это значит , что гормон инсулин больше не производится. Это вызывает сахарный диабет I типа (аутоиммунный диабет). Затем необходимо регулярно вводить инсулин или его аналоги путем подкожной инъекции, в зависимости от результата измерения глюкометром .
Приобретенный диабет
Нарушение регуляции уровня глюкозы называется промежуточной гипергликемией, а в более тяжелых формах - сахарным диабетом II типа (синоним приобретенного диабета , инсулинорезистентности ). Повторяющийся или постоянный высокий уровень сахара в крови обычно указывает на сахарный диабет. В зависимости от степени тяжести могут применяться пероральные противодиабетические препараты . Регулярные упражнения и избегание ожирения снижают риск сахарного диабета II типа, а также упражнения рекомендуются для лечения сахарного диабета II типа.
Избыточный вес и жирная печень
Кроме того, повышенное потребление глюкозы приводит к ожирению и, как следствие, частично к метаболическому синдрому с неалкогольным жировым гепатитом печени , но не к потреблению глюкозы в рамках нормального потребления калорий.
Аналитика
В частности, когда молекула глюкозы быть обнаружены в определенном положении в более крупной молекулы, А спектроскопия ядерного магнитного резонанса , анализ кристаллической структуры рентгеновских лучей или лектин - иммунным окрашиванием с конканавалином А - фермент - репортер - конъюгат (только связывает глюкозу или манноза).
Классические качественные реакции обнаружения
Эти реакции имеют только историческое значение:
Реакция Фелинга
Образец Фелинга является классическим доказательством альдозы. Из-за мутаротации небольшая часть глюкозы всегда присутствует в виде альдегида с открытой цепью. При добавлении реагентов Фелинга (раствор Фелинга (I) и раствор Фелинга (II)) альдегидная группа окисляется до карбоновой кислоты, в то время как комплекс винной кислоты Cu 2+ восстанавливается до Cu + и в виде осадка кирпично-красного цвета (Cu 2 O) не работает.
Отличная реакция
В образце Толленса после добавления аммиачного AgNO 3 к раствору образца Ag + восстанавливается с глюкозы до элементарного серебра.
Образец Barfoedsche
В образце Barfoed в смесь растворенного ацетата меди , ацетата натрия и уксусной кислоты смешивают с раствором сахара , чтобы изучить и нагревали на водяной бане в течение нескольких минут. Глюкоза и другие моносахариды быстро приобретают красноватый цвет и красновато-коричневый оксид меди (I) (Cu 2 O).
Реактив Ниландера
В качестве редуцирующего сахара глюкоза вступает в реакцию с реактивом Ниландера .
Дополнительные доказательства
Когда разбавленный раствор гидроксида калия с глюкозой нагревается до 100 ° C, появляется сильное красноватое потемнение и карамельный запах. Концентрированная серная кислота растворяет сухую глюкозу без почернения при комнатной температуре с образованием сахарной серной кислоты . В растворе с дрожжами при спиртовой ферментации сразу образуется углекислый газ в соотношении 2,0454 молекулы глюкозы на одну молекулу CO 2 . Глюкоза образует с хлоридом олова черную массу. В аммиачном растворе серебра глюкоза (как лактоза и декстрин ) приводит к осаждению серебра без образования серебряного зеркала . В аммиачном растворе ацетата свинца белый гликозат свинца образуется в присутствии глюкозы , которая становится менее растворимой и становится коричневой при приготовлении. В аммиачном растворе меди желтый гидрат оксида меди образуется с глюкозой при комнатной температуре , а при кипячении, с другой стороны, образуется красный оксид меди (также с декстрином, за исключением аммиачного раствора ацетата меди). С реактивом Хагера глюкоза при приготовлении образует оксид ртути . В щелочном растворе висмута элементарный черно-коричневый висмут откладывается с глюкозой. Кипяченная в растворе молибдата аммония глюкоза окрашивает раствор в голубой цвет. Раствор с индигокармином и карбонатом натрия меняет цвет при кипячении с глюкозой.
Инструментальное количественное определение
Рефрактометрия и поляриметрия
В концентрированных растворах глюкозы с небольшим количеством других углеводов их концентрацию можно определить с помощью поляриметра . В случае сахарных смесей концентрацию можно определить с помощью рефрактометра , например, при определении Оксле в процессе производства вина .
Фотометрическо-ферментативный метод в растворе
Фермент глюкозооксидаза (GOx) превращает глюкозу в глюконовую кислоту и перекись водорода с помощью кислорода . Другой фермент, пероксидаза, катализирует хромогенную реакцию (реакция Триндера) фенола с 4-аминоантипирином с образованием пурпурного красителя.
Фотометрический метод тест-полосок
В методе тест-полосок используется упомянутое выше ферментативное превращение глюкозы в глюконовую кислоту с образованием перекиси водорода. Реагенты иммобилизованы на полимерной матрице, так называемой тест-полоске, которая приобретает более или менее яркий цвет. Это можно считать рефлектометрическим методом с помощью портативного светодиодного фотометра на длине волны 510 нм. Это позволяет непрофессионалам проводить рутинное тестирование уровня глюкозы в крови. В дополнение к реакции фенола с 4-аминоантипирином были разработаны новые хромогенные реакции, которые позволяют проводить фотометрию на более высоких длинах волн (550 нм, 750 нм).
Амперометрические датчики глюкозы
Электроанализ глюкозы также основан на ферментативной конверсии , упомянутую выше. Полученный пероксид водорода можно количественно определить амперометрически путем анодного окисления при потенциале 600 мВ. GOx иммобилизован на поверхности электрода или в мембране, расположенной рядом с электродом. Помимо классических драгоценных металлов, таких как платина или золото, в последнее время все чаще используются электроды из углеродных нанотрубок. Б. были легированы бором. Нанопроволоки Cu-CuO также используются в качестве безферментных амперометрических электродов. Таким образом был достигнут предел обнаружения 50 мкмоль / л. Особенно многообещающим методом является так называемая «ферментная разводка». Электроны, протекающие во время окисления, отводятся непосредственно от фермента к электроду через молекулярную проволоку.
Другие сенсорные методы
Есть ряд других химических сенсоров глюкозы. Ввиду важности анализа глюкозы в биологических науках, также были разработаны многочисленные оптические зонды для сахаридов, основанные на использовании бороновых кислот, которые особенно подходят для внутриклеточных сенсорных приложений, где другие (оптические) методы вообще отсутствуют. или может использоваться только в ограниченной степени. В дополнение к производным органической бороновой кислоты, которые часто очень специфично связываются с 1,2-диольными группами сахара, существуют другие концепции зондов, классифицированные в соответствии с функциональными механизмами, которые используют селективные связывающие глюкозу белки (например, конканавалин A) в качестве рецепторов. . Кроме того, были разработаны методы, которые измеряют концентрацию глюкозы косвенно через концентрацию продуктов метаболизма, например Б. потребление кислорода с помощью люминесцентных оптических датчиков. Наконец, существуют концепции, основанные на ферментах, которые используют собственное поглощение или флуоресценцию (меченных флуоресценцией) ферментов в качестве носителей информации.
Иодометрия меди
Глюкозу можно получить из меди - можно определить иодометрию .
Хроматографический процесс
Специально для анализа сложных смесей, содержащих глюкозу, например. Б. В меде сегодня используются хроматографические методы, такие как высокоэффективная жидкостная хроматография и газовая хроматография , часто в сочетании с масс-спектрометрией . Принимая во внимание соотношение изотопов , этот анализ также может надежно обнаружить фальсификацию меда, вызванную добавлением сахара. Получение производных с использованием силилирующих реагентов оказалось выгодным . Таким образом можно также количественно определить пропорции ди- и трисахаридов .
Анализ in vivo
Поглощение глюкозы клетками организмов исследуют с помощью 2-дезокси- D- глюкозы или фтордезоксиглюкозы . ( 18 F) -Фтордезоксиглюкоза используется в онкологии и неврологии в качестве индикатора в позитронно-эмиссионной томографии , где она является наиболее широко используемым диагностическим инструментом.
литература
- Йохен Леманн: Углеводы. Химия и биология. 2., переделка. и эксп. Версия. Тиме, Штутгарт / Нью-Йорк 1996, ISBN 3-13-532902-X .
- Ганс Фогель: таблицы сахаров и их производных. Springer-Verlag, Берлин 2013, ISBN 978-3-642-47764-5 .
- Гюнтер Вольф: метаболизм сахара - биографическое исследование. В кн . : Медицинские ежемесячники . Том 12, 1958, стр. 766-774 и 838-846.
веб ссылки
Индивидуальные доказательства
- ↑ Вход на ГЛЮКОЗАХ в CosIng базе данных Комиссии ЕС, доступ к 16 февраля 2020 года.
- ↑ a b c d Запись для № CAS. 50-99-7 в базе вещества GESTIS из в МРС , доступ к 18 сентября 2014 года. (Требуется JavaScript)
- ↑ Альфред Тёпель: Химия и физика молока: Натуральный материал - Сырье - Пища . Behr's Verlag DE, 2004, ISBN 3-89947-131-8 , стр. 101 ( ограниченный предварительный просмотр в поиске Google Книг).
- ↑ Дэвид Р. Лид (Ред.): Справочник по химии и физике CRC . 90-е издание. (Интернет-версия: 2010 г.), CRC Press / Taylor and Francis, Boca Raton, FL, Physical Constants of Organic Compounds, pp. 3-268.
- ↑ a b c Запись о d-глюкозе. В: Römpp Online . Георг Тиме Верлаг, по состоянию на 31 мая 2013 г.
- ↑ Лист данных по глюкозе (PDF) от Карла Рота , по состоянию на 24 августа 2010 г.
- ↑ Тенар, Гей-Люссак, Био, Дюма: Раппорт о воспоминаниях М. Пелиджио, intitulé: Recherches sur la nature et les propriétés chimiques des Sucres . В: Comptes rendus , 1838, том 7, стр. 106-113.
- ↑ Абрахам Дж. Домб, Джозеф Кост, Дэвид Уайзман: Справочник по биоразлагаемым полимерам . CRC Press, 1998. ISBN 978-1-4200-4936-7 . С. 275.
- ↑ Кенджи Камиде: Целлюлоза и производные целлюлозы . Elsevier, 2005, ISBN 978-0-08-045444-3 , стр. 1.
- ^ Примерный список ВОЗ основных лекарственных средств. (PDF) (Больше не доступно в Интернете.) В: Всемирная организация здравоохранения. Октябрь 2013, архивируются с оригинала апреля 23, 2014 ; Доступ к 22 апреля 2014 года .
- ^ A b Бенджамин Кабальеро, Пол Финглас, Фидель Толдра: Энциклопедия еды и здоровья . Академическая пресса (2016). ISBN 978-0-12-384953-3 , том 3, стр. 239, 608.
- ↑ Маргграф: Опыт химикатов, совершенных в данс ле Dessein de Tirer, un true sucre de разнообразных растений, qui croissent dans nos contrées. «Химические эксперименты с целью получения настоящего сахара из различных растений, произрастающих на наших землях». В: Histoire de l'académie royale des Sciences et belles-lettres de Berlin (1747), стр. 90: Les raisins secs, etant humectés d'une petite Quantité d'eau, de maniere qu'ils mollissent, peuvent alors etre pilés , & le Sucqu'on en exprime, etant depuré & épaissi, fournira une espece de Sucre. «Изюм, смоченный в небольшом количестве воды, чтобы он стал мягче, можно отжимать, а из отжатого сока, очищенного и загущенного, получается своего рода сахар».
- ↑ Аарон Дж. Айде: Развитие современной химии. Харпер и Роу, Нью-Йорк, 1964.
- ↑ a b Джон Ф. Робит: Основы химии углеводов. (= Springer Advanced Texts in Chemistry ) 1998, ISBN 0-387-94951-8 .
- ↑ Предварительные правила для номенклатуры углеводов, Журнал биологической химии, № 247, стр. 613, 1972.
- ↑ Джон Ф. Робит: Основы химии углеводов. Springer Science & Business Media, 2012, ISBN 978-1-4612-1622-3 . С. 7.
- ↑ М.А. Розанов: О классификации стереоизомеров Фишера. В: Журнал Американского химического общества . 28, 1906, с. 114, DOI: 10.1021 / ja01967a014 .
- ^ Эмиль Фишер - Биографический. В: nobelprize.org. 15 июля 1919, доступ к 4 июня 2018 .
- ↑ Берт Фрейзер-Рид: глюкоза Ван 'т Гоффа . In: Chem. Eng. Новости , 1999 г., том 77, выпуск 39, стр. 8; DOI: 10.1021 / cen-v077n039.p008 .
- ^ IUPAC: Номенклатура углеводов (Рекомендации 1996 г.) .
- ↑ Отто Мейерхоф - Факты. В: nobelprize.org. 15 июля 2018, доступ к 15 июля 2018 .
- ^ Ганс фон Эйлер-Челпин - Факты. В: nobelprize.org. Проверено 3 сентября 2018 года .
- ↑ Артур Харден - Факты. В: nobelprize.org. 17 июня 1940, доступ к 3 сентября 2018 .
- ↑ Бернардо Хусей - Факты. В: nobelprize.org. 21 сентября 1971, доступ к 15 июля 2018 .
- ^ Карл Кори - Факты. В: nobelprize.org. 20 октября 1984, доступ к 15 июля 2018 .
- ^ Герти Кори - Факты. В: nobelprize.org. 26 октября 1957, доступ к 15 июля 2018 .
- ↑ Луис Лелуар - Факты. В: nobelprize.org. 15 июля 2018, доступ к 15 июля 2018 .
- ↑ Фред В. Шенк: Глюкоза и глюкозосодержащие сиропы . В: Энциклопедия промышленной химии Ульмана , Wiley-VCH, Weinheim, 2006. doi : 10.1002 / 14356007.a12_457.pub2 .
- ↑ Патрик Ф. Фокс: Advanced Dairy Chemistry Том 3: Лактоза, вода, соли и витамины , Springer, 1992. Том 3, ISBN 978-0-412-63020-0 . С. 316.
- ^ Бенджамин Кабальеро, Пол Финглас, Фидель Толдра: Энциклопедия еды и здоровья . Академическая пресса (2016). ISBN 978-0-12-384953-3 , том 1, стр. 76.
- ↑ Х. Ф. Банн, П. Дж. Хиггинс: Реакция моносахаридов с белками: возможное эволюционное значение . В кн . : Наука . 213, No. 4504, 1981, pp. 222-224. DOI : 10.1126 / science.12192669 .
- ↑ Джереми М. Берг: Биохимия Страйера. Springer-Verlag, 2017, ISBN 978-3-662-54620-8 , стр. 531.
- ^ Реджинальд Х. Гарретт: Биохимия. Обучение Cengage, 2012, ISBN 978-1-133-10629-6 . С. 194, 199.
- ^ Дональд Воет, Джудит Г. Воет: Биохимия, 4-е издание. Джон Вили и сыновья, 2010, ISBN 978-0-470-57095-1 . С. 363.
- ↑ Альберт Л. Ленингер, Биохимия, 6-е издание , Worth Publishers Inc. 1972, ISBN 0-87901-009-6 , стр. 228.
- ↑ a b c Питер К. Генрих: Биохимия и патобиохимия Леффлера / Петридеса. Springer-Verlag, 2014, ISBN 978-3-642-17972-3 , стр. 27.
- ↑ Эусебио Хуаристи, Габриэль Куэвас: The Anomeric Effect CRC Press, 1995. ISBN 0-8493-8941-0 . С. 9-10.
- ↑ a b Манфред Гессе, Герберт Мейер, Бернд Зе, Стефан Биенц, Лоран Биглер, Томас Фокс: Спектроскопические методы в органической химии . 8., перераб. Версия. Георг Тиме, 2011, ISBN 978-3-13-160038-7 , стр. 34 .
- ^ Базы данных о составе продуктов питания Показать список продуктов. В: ndb.nal.usda.gov. Проверено 25 августа 2018 года .
- ^ Сахар . ( Сувенир от 18 июля 2011 г. в Интернет-архиве ) Государственный университет штата Орегон ; доступ 28 июня 2018 г.
- ↑ a b c d e f g h i P J Fellows: Food Processing Technology. Издательство Woodhead, 2016 г., ISBN 978-0-08-100523-1 , стр.197.
- ↑ a b Томас Беккер, Дитмар Брайтхаупт, Хорст Вернер Доэлле, Армин Фихтер, Гюнтер Шлегель, Сакаю Симидзу, Хидеаки Ямада: биотехнологии . В: Энциклопедия промышленной химии Ульмана . 7-е издание. Wiley-VCH, 2011, ISBN 978-3-527-32943-4 , том 6, с. 48.
- ↑ Крахмальный сахар . В: Warenlexikon компании Merck . 3-е изд. 1884 и сл., С. 457 ф.
- ^ А б Алан Дэвидсон: Оксфордский компаньон к еде. ОУП Оксфорд, 2014 г., ISBN 978-0-19-104072-6 , стр. 527.
- ↑ a b c Японское общество исследования амилазы: Справочник по амилазам и родственным ферментам. Эльзевир, 2014, ISBN 978-1-4832-9939-6 , стр.195.
- ^ Г.Б. Мэдсен, Б.Е. Норман, С. Слотт: Новая термостойкая бактериальная амилаза и ее использование при высокотемпературном сжижении . В: Крахмал (1973), том 25, выпуск 9, DOI: 10.1002 / star.19730250906 . С. 304-308.
- ^ BE Norman: новый фермент разветвления для применения в промышленности сиропа глюкозы. В: Крахмал - сила , 34, 1982, с. 340, DOI: 10.1002 / star.19820341005 .
- ↑ Джеймс Н. Бемиллер, Рой Л. Уистлер: Крахмал: химия и технология (= пищевая наука и технология), 3-е. Издание, Academic Press, Нью-Йорк, 2009 г., ISBN 0-08-092655-X .
- ↑ Кукурузный сироп с высоким содержанием фруктозы: вопросы и ответы. (Больше не доступны в Интернет.) США пищевых продукты и медикаменты, 5 ноября 2014, в архиве от оригинала Января 25, 2018 ; Доступ к 18 декабря 2017 года .
- ↑ Кевин Панг: « Мексиканская кока-кола» стала хитом в США ( воспоминание от 29 июня 2011 г. в Интернет-архиве ) В: « Сиэтл Таймс» от 29 октября 2004 г.
- ↑ Стив Т. Беккет: Промышленное производство и использование шоколада Беккета. John Wiley & Sons, 2017, ISBN 978-1-118-78014-5 , стр. 82.
- ^ Ханс-Дитер Белиц , Вернер Грош , Петер Шиберле : пищевая химия . Springer, Berlin 2009. ISBN 978-3-540-69935-4 . С. 270-289.
- ^ Натан Мирвольд , Крис Янг, Максим Билет: Модернистская кухня: искусство и наука кулинарии . Кулинарная лаборатория 2011. ISBN 978-0-9827610-0-7 . Том 3, стр. 89 и далее.
- ↑ Джеймс А. Кент: Справочник Ригеля по промышленной химии. Springer Science & Business Media, 2013, ISBN 978-1-4757-6431-4 , стр. 938.
- ↑ Ашок Пандей: Промышленные биоперерабатывающие заводы и Белая биотехнология. Эльзевир, 2015, ISBN 978-0-444-63464-1 , стр. 488.
- ↑ a b c Питер К. Генрих: Биохимия и патобиохимия Леффлера / Петридеса. Springer-Verlag, 2014, ISBN 978-3-642-17972-3 , стр.195.
- ↑ а б в г д У. Сатьянараяна: Биохимия. Elsevier Health Sciences, 2014, ISBN 978-81-312-3713-7 . С. 674.
- ↑ Д.Х. Вассерман: Четыре грамма глюкозы. В: Американский журнал физиологии - эндокринология и метаболизм. Том 296, номер 1, январь 2009 г., стр. E11-E21, doi: 10.1152 / ajpendo.90563.2008 , PMID 18840763 , PMC 2636990 (полный текст).
- ↑ б с д е е г Peter C. Heinrich: Löffler / Петридес биохимии и Pathobiochemistry. Springer-Verlag, 2014, ISBN 978-3-642-17972-3 , стр. 199, 200.
- ^ Б Peter C. Heinrich: LÖFFLER / Петридес биохимия и Pathobiochemistry. Springer-Verlag, 2014, ISBN 978-3-642-17972-3 , стр. 214.
- ↑ Основы гликобиологии , Аджит Варки (ред.), 2-е. Издание, издательство Cold Spring Harbor Laboratories Press , ISBN 978-0-87969-770-9 . Архивировано из оригинального 6 -го декабря 2016 года.
- ↑ Питер К. Генрих: Löffler / Petrides Biochemistry and Pathobiochemistry . Springer-Verlag, 2014, ISBN 978-3-642-17972-3 , стр.404.
- ↑ Результат поиска UniProt .
- ↑ Результат поиска UniProt .
- ↑ Гарольд А. Харпер: Медицинская биохимия. Springer-Verlag, 2013, ISBN 978-3-662-22150-1 , стр. 641.
- ↑ AM Navale, AN Paranjape: Транспортеры глюкозы: физиологические и патологические роли. В: Биофизические обзоры . Том 8, номер 1, март 2016 г., стр. 5-9, doi: 10.1007 / s12551-015-0186-2 , PMID 28510148 , PMC 5425736 (полный текст).
- ↑ Б. Торенс: GLUT2, определение глюкозы и гомеостаз глюкозы. В кн . : Диабетология . Том 58, номер 2, февраль 2015 г., стр. 221-232, DOI : 10.1007 / s00125-014-3451-1 , PMID 25421524 .
- ^ RC Bonadonna, S. Del Prato, E. Bonora, MP Saccomani, G. Gulli, A. Natali, S. Frascerra, N. Pecori, E. Ferrannini, D. Bier, C. Cobelli, RA DeFronzo: Роли глюкозы транспорт и фосфорилирование глюкозы при мышечной инсулинорезистентности NIDDM. В: Диабет. Том 45, номер 7, июль 1996 г., стр. 915-925, PMID 8666143 .
- ↑ С. Хуанг, член парламента Чехии: Транспортер глюкозы GLUT4. В кн . : Клеточный метаболизм . Том 5, номер 4, апрель 2007 г., стр. 237-252, DOI : 10.1016 / j.cmet.2007.03.006 , PMID 17403369 .
- ↑ Р. Говерс: Клеточная регуляция поглощения глюкозы переносчиком глюкозы GLUT4. В кн . : Успехи клинической химии . Том 66, 2014, стр. 173-240, PMID 25344989 .
- ↑ C. Ghezzi, DD Loo, EM Wright: Физиология почечной обработки глюкозы через SGLT1, SGLT2 и GLUT2. В кн . : Диабетология . [Электронная публикация перед отправкой в печать] Август 2018 г., DOI: 10.1007 / s00125-018-4656-5 , PMID 30132032 .
- ↑ С.Б. Поульсен, Р.А. Фентон, Т. Риг: Котранспорт натрия и глюкозы. В: Современное мнение в нефрологии и артериальной гипертонии. Том 24, номер 5, сентябрь 2015 г., стр. 463-469, doi: 10.1097 / MNH.0000000000000152 , PMID 26125647 , PMC 5364028 (полный текст).
- ^ Дональд Воет, Джудит Г. Воет: Биохимия, 4-е издание . Джон Вили и сыновья, 2010, ISBN 978-0-470-57095-1 . С. 359.
- ^ A b Дональд Воет, Джудит Г. Воет: Биохимия, 4-е издание. Джон Вили и сыновья, 2010, ISBN 978-0-470-57095-1 . С. 59.
- ↑ a b Leszek Szablewski: Гомеостаз глюкозы и инсулинорезистентность. Издательство Bentham Science, 2011, ISBN 978-1-60805-189-2 , стр. 46.
- ↑ AM Smith, SC Zeeman, SM Smith: Распад крахмала. В: Ежегодный обзор биологии растений . Том 56, 2005 г., стр. 73-98, DOI: 10.1146 / annurev.arplant.56.032604.144257 , PMID 15862090 .
- ↑ MM-Adeva any tax, N. Pérez-Felpete, C. Fernandez-Fernandez, C. Donapetry-García, C. Pazos-García: метаболизм глюкозы в печени у человека. В: Bioscience Reports . Volume 36, number 6, 12 2016, p. E00416, doi: 10.1042 / BSR20160385 , PMID 27707936 , PMC 5293555 (полный текст).
- ^ Х. Роберт Хортон, Лоуренс А. Моран, К. Грей Скримджер, Марк Д. Перри, Дж. Дэвид Рон и Карстен Биле (переводчик): Biochemie . Исследования Пирсона; 4-е обновленное издание 2008 г .; ISBN 978-3-8273-7312-0 ; С. 490-496.
- ^ Б Brian K. Hall: Evolution Strickberger в. Jones & Bartlett Publishers, 2013, ISBN 978-1-4496-1484-3 , стр. 164.
- ^ Реджинальд Х. Гарретт: Биохимия. Обучение Cengage, 2012 г., ISBN 978-1-133-10629-6 , стр. 551.
- ↑ Дж. Дж. Джонс: Глюкоза в печени и метаболизм липидов. В кн . : Диабетология . Том 59, номер 6, 06 2016 г., стр. 1098–1103, DOI: 10.1007 / s00125-016-3940-5 , PMID 27048250 .
- ↑ Н. Энтнер, М. Дудоров: Окисление глюкозы и глюконовой кислоты Pseudomonas saccharophila . В: J Biol Chem . , 1952, 196 (2), pp. 853-862 (англ.); PMID 12981024 ; jbc.org (PDF).
- ^ А. Аннибальди, К. Видманн: метаболизм глюкозы в раковых клетках. В: Current Opinion in Clinical Nutrition and Metabolic Care . Том 13, номер 4, июль 2010 г., стр. 466-470, DOI : 10.1097 / MCO.0b013e32833a5577 , PMID 20473153 .
- ↑ XD Xu, SX Shao, HP Jiang, YW Cao, YH Wang, XC Yang, YL Wang, XS Wang, HT Niu: эффект Варбурга или обратный эффект Варбурга? Обзор метаболизма рака. В кн . : Исследования и лечение онкологии . Том 38, номер 3, 2015 г., стр. 117-122, DOI: 10.1159 / 000375435 , PMID 25792083 .
- ↑ Л. Саблевски: Экспрессия переносчиков глюкозы при раке. В: Biochimica et Biophysica Acta . Том 1835, номер 2, апрель 2013 г., стр. 164-169, DOI : 10.1016 / j.bbcan.2012.12.004 , PMID 23266512 .
- ↑ К. Адекола, С. Т. Розен, М. Шанмугам: Транспортеры глюкозы в метаболизме рака. В: Современное мнение в онкологии . Том 24, номер 6, ноябрь 2012 г., стр. 650-654, DOI : 10.1097 / CCO.0b013e328356da72 , PMID 22913968 .
- ^ RH Де Декен: Эффект Крэбтри: регуляторная система в дрожжах. В: Журнал общей микробиологии . Том 44, номер 2, август 1966 г., стр. 149-156, DOI : 10.1099 / 00221287-44-2-149 , PMID 5969497 .
- ↑ Э. де Олд Риис, Ф. Картени, П. Параскандола, Дж. Серпа, С. Маццолени: Пересмотр эффекта Крэбтри / Варбурга в динамической перспективе: фитнес-преимущество против гибели клеток, вызванной сахаром. В кн . : Cell Cycle . Volume 17, number 6, 2018, pp. 688-701, doi: 10.1080 / 15384101.2018.1442622 , PMID 29509056 , PMC 5969562 (полный текст).
- ↑ Георг Шведт Цукерсус, химия. John Wiley & Sons, 2012, ISBN 978-3-527-66001-8 , стр. 100.
- ↑ Шмидт, Ланг: Physiologie des Menschen, 30-е издание. Springer Verlag, 2007, с. 907.
- ↑ T. Dandekar, S. Schuster, B. Snel, M. Huynen, P. Bork: Pathway alignment: приложение к сравнительному анализу гликолитических ферментов. В кн . : Биохим. J. 343 Pt 1, 1999, стр. 115-124 ( PMID 10493919 ; PMC 1220531 (свободный полный текст)).
- ↑ а б в Л. Л. Куккук, Дж. Д. Мул, С. Э. ла Флер: Определение уровня глюкозы в системе вознаграждения. В: Границы неврологии . Volume 11, 2017, p. 716, doi: 10.3389 / fnins.2017.00716 , PMID 29311793 , PMC 5742113 (полный текст).
- ↑ a b Р. М. Такер, С. Ю. Тан: Влияют ли непитательные подсластители на острый гомеостаз глюкозы у людей? Систематический обзор. В кн . : Физиология и поведение . Том 182, декабрь 2017 г., стр. 17-26, DOI: 10.1016 / j.physbeh.2017.09.016 , PMID 28939430 .
- ^ SE La Fleur, E. Fliers, A. Kalsbeek: Неврология гомеостаза глюкозы. В кн . : Справочник по клинической неврологии . Том 126, 2014 г., стр. 341-351, DOI: 10.1016 / B978-0-444-53480-4.00026-6 , PMID 25410233 .
- ^ PH Bisschop, E. Fliers, A. Kalsbeek: вегетативная регуляция производства глюкозы в печени. В кн . : Комплексная физиология . Том 5, номер 1, январь 2015 г., стр. 147-165, DOI : 10.1002 / cphy.c140009 , PMID 25589267 .
- ^ WA Scherbaum, BM Lobnig, In: Hans-Peter Wolff, Thomas R. Weihrauch: Внутренняя терапия 2006, 2007. 16-е издание. Эльзевир, Мюнхен, 2006 г., ISBN 3-437-23182-0 , стр. 927, 985.
- ↑ Гарольд А. Харпер: Медицинская биохимия. Springer-Verlag, 2013, ISBN 978-3-662-22150-1 , стр. 294.
- ↑ а б в Донард Дуайер: метаболизм глюкозы в головном мозге. Academic Press, 2002, ISBN 978-0-12-366852-3 , стр. XIII.
- ↑ SH Fairclough, K. Houston: Метаболическая мера умственного усилия. В кн . : Биологическая психология . Том 66, номер 2, апрель 2004 г., стр. 177-190, DOI : 10.1016 / j.biopsycho.2003.10.001 , PMID 15041139 .
- ↑ MT Gailliot, RF Baumeister: Физиология силы воли: связь уровня глюкозы в крови с самоконтролем. В: Обзор личности и социальной психологии: официальный журнал Общества личности и социальной психологии, Inc. Том 11, номер 4, ноябрь 2007 г., стр. 303-327, DOI : 10.1177 / 1088868307303030 , PMID 18453466 .
- ^ A b Ричард А. Харви, Дениз Р. Ферье: Биохимия. 5-е издание, Lippincott Williams & Wilkins, 2011, ISBN 978-1-60831-412-6 , стр. 366.
- ↑ а б У. Сатьянараяна: Биохимия. Elsevier Health Sciences, 2014, ISBN 978-81-312-3713-7 , стр. 508.
- ↑ Ш.Холт, Дж. Миллер, П. Петоч: Индекс инсулина в продуктах питания: потребность в инсулине , вызванная порциями обычных продуктов по 1000 кДж. В: Американский журнал клинического питания . Том 66, выпуск 5, ноябрь 1997 г., стр. 1264-1276, DOI: 10.1093 / ajcn / 66.5.1264 , PMID 9356547 .
- ^ П. Конканнон, С. С. Рич, Г. Т. Непом: Генетика диабета типа 1А. В: Медицинский журнал Новой Англии . Volume 360, Number 16, April 2009, pp. 1646-1654, DOI: 10.1056 / NEJMra0808284 , PMID 19369670 .
- ↑ Р. Гоял, И. Джиалал: Непереносимость глюкозы. В: StatPearls [Интернет]. Остров сокровищ (Флорида), 2018. PMID 29763085 .
- ↑ Р. Буреш: Физические упражнения и контроль уровня глюкозы. В кн . : Журнал спортивной медицины и физической подготовки . Volume 54, Number 4, August 2014, pp. 373-382, PMID 25034542 .
- ↑ Х. Ики-Ярвинен: Неалкогольная жировая болезнь печени как причина и следствие метаболического синдрома. В кн . : Ланцет . Диабет и эндокринология. Том 2, номер 11, ноябрь 2014 г., стр. 901-910, DOI : 10.1016 / S2213-8587 (14) 70032-4 , PMID 24731669 .
- ↑ Х. Фелинг: Количественное определение сахара в моче. В: Архивы физиологической медицины (1848 г.), том 7, стр. 64–73.
- ↑ Б. Толленс: О растворе аммиачного серебра как реагенте на альдегид . В: Отчеты Немецкого химического общества . 15, 1882, стр. 1635-1639.
- ↑ К. Барфоед: Об обнаружении виноградного сахара в дополнение к декстрину и связанным с ним телам . В кн . : Журнал аналитической химии . 12, No. 1, 1873, p. 27. doi : 10.1007 / BF01462957 .
- ^ Эмиль Ниландер: О щелочном растворе висмута как реагенте для определения глюкозы в моче , журнал по физиологической химии . Volume 8, Issue 3, 1884, pp. 175-185 ( аннотация ).
- ↑ б с д е е г ч я J K Георг Шведта: Zuckersweet химии. Джон Вили и сыновья, 2012 г., ISBN 978-3-527-66001-8 , стр. 102.
- ↑ П. Триндер: Определение глюкозы в крови с помощью глюкозооксидазы с альтернативным акцептором кислорода. В кн . : Анналы клинической биохимии . 6, 1969, стр. 24–27; DOI: 10.1177 / 000456326900600108 .
- ↑ М. Мизогучи, М. Исияма, М. Сига, К. Сасамото: Водорастворимый хромогенный реагент для колориметрического определения перекиси водорода - альтернатива 4-аминоантипирину, работающему на длинных волнах. В кн . : Аналитические коммуникации . 35, 1998, стр. 71-73; DOI: 10.1039 / A709038B .
- ↑ Дж. Ван: Электрохимические биосенсоры глюкозы. В: Chem. Rev. 108, 2008, стр. 814-825; PMID 18154363 .
- ↑ X. Chen, J. Chen, Ch. Deng, Ch. Xiao, Y. Yang, Z. Nie, S. Yao: Амперометрический биосенсор глюкозы на основе электрода, модифицированного углеродными нанотрубками, легированными бором. В кн . : Таланта . 76, 2008, стр. 763-767; DOI: 10.1016 / j.talanta.2008.04.023 ; PMID 18656655 .
- ↑ Г. Ван, Ю. Вэй, В. Чжан, Х. Чжан, Б. Фанг, Л. Ван: Безферментное амперометрическое определение глюкозы с использованием композитных нанопроволок Cu-CuO. В: Microchimica Acta . 168, 2010, стр. 87–92; DOI: 10.1007 / s00604-009-0260-1 .
- ^ Т. Дж. Охара, Р. Раджагопаян, А. Хеллер: «Проводные» ферментные электроды для амперометрического определения глюкозы или лактата в присутствии мешающих веществ. В: Анал. Chem. , 66, 1994, стр 2451-2457. DOI: 10.1021 / ac00087a008 ; PMID 8092486 .
- ↑ a b c Борисов С. М., Вольфбейс О. С. Оптические биосенсоры. В: Chem. Rev. 108, 2008, стр. 423-461; DOI: 10,1021 / cr068105t ; PMID 18229952 .
- ↑ С. Ферри, К. Кодзима, К. Соде: Обзор глюкозооксидаз и глюкозодегидрогеназ: взгляд на ферменты, чувствительные к глюкозе, с высоты птичьего полета. В: Журнал диабетической науки и техники . Том 5, номер 5, сентябрь 2011 г., стр. 1068-1076, DOI: 10.1177 / 193229681100500507 , PMID 22027299 , PMC 3208862 (полный текст).
- ↑ HS Mader, OS Wolfbeis: Зонды на основе бороновой кислоты для микроопределения сахаридов и гликозилированных биомолекул. В: Microchimica Acta , 162, 2008, стр. 1-34; DOI: 10.1007 / s00604-008-0947-8 .
- ↑ OS Wolfbeis, I. Oehme, N. Papkovskaya, I. Klimant: Золь-гель глюкозные биосенсоры с оптическими преобразователями кислорода и метод компенсации переменного кислородного фона. В: Биосенсоры и биоэлектроника . 15, 2000, стр. 69–76; DOI: 10.1016 / S0956-5663 (99) 00073-1 .
- ↑ а б А. Л. Галант, Р. К. Кауфман, Дж. Д. Уилсон: Глюкоза: обнаружение и анализ. В кн . : Пищевая химия . Том 188, декабрь 2015 г., стр. 149-160, DOI : 10.1016 / j.foodchem.2015.04.071 , PMID 26041177 .
- ↑ М.Л. Санс, Х. Санс, И. Мартинес-Кастро: Газовый хроматографо-масс-спектрометрический метод качественного и количественного определения дисахаридов и трисахаридов в меде. В: Journal of Chromatography A 1059 (1-2), 2004, стр. 143-148; PMID 15628134 .
- ↑ hummel@mpimp-golm.mpg.de: GMD - Глюкоза (1MEOX) (5TMS) BP - InChI = 1S / C22H55NO6Si5 / c1-24-23-17-19 (26-31 (5.6) 7) 21 (28- 33 (11,12) 13) 22 (29-34 (14,15) 16) 20 (27-32 (8,9) 10) 18-25-30 (2,3) 4 / ч 17,19- 22H. В: gmd.mpimp-golm.mpg.de. 19 июля 2007. Проверено 4 июня 2018 .
- ^ AI Cabañero, JL Recio, M. Rupérez: Жидкостная хроматография в сочетании с масс-спектрометрией изотопного соотношения: новый взгляд на обнаружение взрослых особей меда. В: J. Agric Food Chem . 54 (26), 27 декабря 2006 г., стр. 9719-9727; PMID 17177492 .
- ↑ М. Беккер, Ф. Лер, Т. Розенау, А. Поттхаст: Этоксиматорно-силилированный подход для анализа моно- и дисахаридов и характеристики их идентификационных параметров с помощью ГХ / МС. В кн . : Таланта . 115, 2013, стр. 642–651; PMID 24054643 .
- ↑ Общество немецких химиков: приложения к позиционному документу Секции ядерной химии ( памятная записка от 31 марта 2010 г. в Интернет-архиве ), февраль 2000 г.
- ↑ Симона Машауэр, Олаф Пранте: Подслащивание фармацевтической радиохимии с помощью 18 F-фторгликозилирования: краткий обзор . В: BioMed Research International , том 2014, ID статьи 214748; DOI: 10.1155 / 2014/214748 ; PMID 24991541 ; PMC 4058687 (полный текст).